Dettagli
- Caratteristiche anatomiche macroscopiche
- Caratteristiche anatomiche microscopiche
- FISIOLOGIA CUTANEA
- Cheratinizzazione
- Funzione protettiva
- Funzione sensoriale
- Funzione secretoria
- Funzione metabolica
- Funzione pigmento-genetica
- Funzione termoregolatrice
- Funzione di assorbimento e di permeabilità selettiva
01.02 – [Introduzione] Struttura generale della cute, lesioni elementari, lesioni microscopiche ed E.O. e terapia topica
Caratteristiche anatomiche macroscopiche
- La cute è l’organo con la superficie più estesa (1,5-2 m2), anche se variabile in rapporto alla massa corporea individuale. Il suo peso medio, nell’adulto, è di circa 5 kg, dato che fa di essa uno degli organi più pesanti.
- Lo spessore della cute è molto variabile in rapporto a età, sesso e sede topografica; per esempio, essa è molto sottile alle palpebre (0,4 mm), mentre è molto spessa sul dorso e sulle superfici palmo-plantari (>3 mm).
- Il colore varia dal bianco-rosato, nei soggetti di etnia nordeuropea, al marrone scuro nei neri africani, in base alle caratteristiche genetiche. Alcune sedi, come le regioni ano-genitali e l’areola mammaria, sono più pigmentate, soprattutto dopo la pubertà. Il colore cutaneo è il risultato della componente vascolare (tonalità rossa), di quella degli xantocromi (tonalità gialla) e della pigmentazione melanica (tonalità bruna).
- La cute è quasi totalmente ricoperta di peli, sottili e corti (peli lanugo o di vello) o di maggiori dimensioni: capelli, sopracciglia, ciglia, tragi (orecchio), vibrisse (narici), peli del pube, peli ascellari. Sono glabre le sedi palmo-plantari e le semimucose.
- La superficie cutanea è solcata da pieghe, legate alla mobilità della pelle a livello articolare (pieghe articolari), alla contrazione dei muscoli sottostanti (pieghe muscolari) e alla diminuzione del pannicolo adiposo e dell’elasticità cutanea (pieghe senili o rughe).
- La pelle è disseminata di piccole depressioni puntiformi centrate dagli osti; questi sono gli orifizi di sbocco dei follicoli piliferi (osti follicolari), delle ghiandole sebacee non annesse ai follicoli e delle ghiandole sudorifere (pori sudoriferi eccrini).
- Sottili solchi lineari, definiti solchi cutanei, congiungono gli osti follicolari vicini, disegnando una rete a maglie losangiche. Particolarmente evidenti sono i solchi cutanei palmari. Altri solchi allineati parallelamente e separati da creste, disposti in modo regolare, sul palmo delle mani e sulla pianta dei piedi, in corrispondenza della superficie palmare dei polpastrelli delle dita hanno una caratteristica configurazione ad anse e a vortice (rosette tattili). Le rosette tattili sono differenti in ogni soggetto e da un dito all’ altro, tanto da costituire un carattere individuale e un sicuro segno di identificazione (dermatoglifì) tramite le impronte digitali.
- La principale caratteristica fisica della cute è rappresentata dalla presenza di uno strato epidermico superficiale cheratinizzato (strato corneo), resistente agli insulti esterni (fisici, chimici, biologici) e semipermeabile. I tessuti sottostanti (derma e ipoderma) sono invece strutture di sostegno dell’epidermide, e garantiscono, tra l’altro, l’elasticità e la distensibilità cutanea.
- L’elasticità consiste nella capacità di distendersi della cute in seguito a trazione e di riacquistare successivamente l’assetto primitivo, grazie alla funzione delle fibre elastiche. La distensibilità, ossia la proprietà di allungarsi, è limitata dalle fibre collagene. In età senile aumenta la distensibilità, mentre diminuisce l’elasticità, con formazione di rughe, borse e cute lassa.
Caratteristiche anatomiche microscopiche
Struttura architetturale
- La pelle è costituita da una struttura superficiale rappresentata da un epitelio pluristratifìcato di origine ectodermica, denominata epidermide, e da una struttura sottostante che ne rappresenta l’ “impalcatura di sostegno”, di natura connettivale, definita derma (di origine mesodermica).
- Le due componenti sono separate da una sottile lamina definita membrana basale (MB) o giunzione dermoepidermica (Immagine 01). Dal punto di vista istologico, al microscopio ottico la MB si evidenzia come un sottile strato positivo alla colorazione PAS (Periodic Acid Schiff, acido periodico di Schiff), mentre alla microscopia elettronica (ME) rivela una maggiore complessità strutturale. La sua porzione superiore è delimitata dal polo inferiore dei cheratinociti dello strato basale epidermico, con i loro emidesmosomi e i rispettivi tonofìlamenti. Al di sotto della membrana cellulare dei cheratinociti dello strato basale inizia quindi la MB, costituita, dall’alto verso il basso, da tre sottili strati sovrapposti: lamina lucida, lamina densa e lamina fìbro-reticolare.
- La lamina lucida appare chiara alla ME ed è attraversata dai filamenti di ancoraggio adesi in corrispondenza degli emidesmosomi delle membrane cellulari dei cheratinociti basali e della sottostante lamina densa. Questo strato contiene la laminina 5, suo principale componente che si lega al collagene di tipo IV e ai proteoglicani; la laminina 5 viene prodotta dai cheratinociti ed è una molecola eterotrimerica costituita da tre catene α3β3ϒ2. Tramite il legame con il recettore integrinico α6β4 tale componente stabilizza la base dei cheratinociti; infatti, nei pazienti con epidermolisi bollosa giunzionale (malattia associata a un difetto del gene che codifica per le catene della laminina) manca l’adesione dei cheratinociti alla MB con il conseguente scollamento epidermico.
- A questo livello, inoltre, si localizza anche l’antigene del pemfìgoide bolloso.
- La lamina densa, così definita per la sua elettrondensità, è costituita prevalentemente da collagene di tipo IV più compatto (legato alla laminina e ai proteoglicani), il quale provvede al supporto strutturale e alla flessibilità.
- La lamina fibra-reticolare, più chiara, è percorsa da fibrille di ancoraggio, costituite da collagene di tipo VII, che si dipartono dalla lamina densa e fanno aderire l’epidermide al derma. In questo strato è localizzato l’antigene dell’epidermolisi bollosa acquisita.
- La più importante funzione della giunzione dermo-epidermica è di garantire un collegamento strutturale tra epidermide e derma e di fornire resistenza contro le forze d’urto esterne. Tale giunzione serve inoltre come supporto all’epidermide, di cui determina la polarità di crescita, condiziona l’organizzazione del citoscheletro delle cellule basali e funziona da barriera semipermeabile.
- Il confine dermo-epidermico delimitato dalla MB non è lineare, ma caratterizzato dalla presenza di ondulazioni, grazie alle digitazioni più o meno allungate del derma superficiale che si insinuano nell’epidermide (papille dermiche). Esse assicurano una perfetta coesione fra l’epitelio e il connettivo sottostante e garantiscono anche una più estesa superficie di contatto e, quindi, di scambi metabolico-funzionali fra i due tessuti.
- L’epidermide è infatti avascolare, mentre il derma è riccamente vascolarizzato da un reticolo vasale superficiale o sottopapillare e da un più sviluppato reticolo vasale profondo dermico, collegati da vasi comunicanti ascendenti. Dal reticolo sub papillare traggono origine i rami terminali capillari siti nelle papille dermiche, estreme propaggini vascolari della cute.
- Al di sotto del derma si ritrova, senza una delimitazione netta, l’ipoderma, costituito da lobuli adiposi separati da tralci connettivali vascolarizzati e innervati. Anche il tessuto adiposo è di origine mesodermica.
- L’ipoderma, in alcune sedi, presenta una componente muscolare liscia (muscoli pellicciai del collo e del viso e muscoli lisci dello scroto, delle piccole e delle grandi labbra).
- Strutture particolari, che hanno la stessa embriogenesi, sono gli annessi cutanei, ossia le ghiandole sudorifere, le ghiandole sebacee, i follicoli piliferi, i peli e le unghie.
Epidermide
- È costituita da un epitelio pluristratificato di elementi cheratinociti o epiteliociti o corneociti), che subiscono una maturazione differenziativa da cui deriva la produzione di cheratina. I cheratinociti sono cellule di derivazione ectodermica contenenti, nel loro citoplasma, filamenti di cheratina che formano, con i cheratinociti adiacenti, desmosomi o giunzioni desmosomiali. I filamenti di cheratina, di cui esistono più di trenta tipi differenti (circa venti epiteliali e dieci dei capelli), rappresentano la componente distintiva dei cheratinociti. Le cheratine si dividono in acide o di tipo 1 (K10-K20) e basiche-neutrali o di tipo 2 (K1-K9). Lo spessore dell’epidermide varia dai 40 µ alle palpebre sino a 1,5 mm e oltre in sede plantare.
- Gli strati cellulari costituenti l’epidermide sono cinque (si veda Immagine 01). Quello a diretto contatto con la MB è lo strato germinativo o basale, composto da un unico insieme di cellule prismatiche a maggior asse verticale, con citoplasma basofilo e nucleo ampio. Il 3-5% delle cellule basali va incontro a continue mitosi, grazie alla presenza di cellule staminali e all’attività della funzione moltiplicativa.
- Il citoplasma delle cellule basali contiene rari tonofìlamenti (costituiti da fibroproteine con gruppi sulfidrilici e ponti disolfuro), che si aggregano in tonofibrille. Queste ultime, assieme a due placche di contatto (ispessimenti delle membrane cellulari di due cheratinociti affiancati) e a una sostanza cementante, costituiscono i desmosomi, sottili ponti che garantiscono la coesione cellulare intraepiteliale e favoriscono gli interscambi tra cheratinociti vicini. L’adesione fra cellule dello strato basale e MB della cute è assicurata dagli emidesmosomi e da filamenti di ancoraggio, attraverso le integrine, le quali costituiscono una famiglia di recettori glicoproteici transmembrana coinvolte nelle interazioni cellula-cellula e cellula-matrice, oltre che nella differenziazione dei cheratinociti.
- La capacità dell’epidermide di mantenere costante il turnover cellulare è basata sulla presenza, nello strato basale e nel follicolo pilifero, di una popolazione di cellule staminali. Queste ultime danno origine sia ad altre cellule staminali sia a cellule committed, ossia che vanno incontro a una differenziazione terminale. Il passaggio da cheratinocita staminale a cheratinocita differenziato avviene attraverso una sottopopolazione di cellule che si trovano in uno stadio maturativo intermedio, le TAC (Transit Amplifyng Cells, cellule di transizione amplificatorie). Le cellule dello strato basale in fase attiva di moltiplicazione mitotica perdono questa loro caratteristica quando si staccano dalla MB e progrediscono verso la superficie cutanea con un processo di differenziazione terminale (cellule committed). Quando raggiungono lo strato granuloso, le cellule iniziano a perdere il nucleo e gli organelli citoplasmatici, grazie a un meccanismo degradativo che comporta l’attivazione parziale del meccanismo dell’ apoptosi; in questo modo i cheratinociti si trasformano nelle squame cheratinizzate dello strato corneo (corneociti). Lo strato spinoso, così definito per la ricca presenza di desmosomi, appendici simili a spine alla periferia dei cheratinociti osservabili in microscopia ottica, a causa delle alterazioni indotte dalla preparazione istologica, è strutturato in piani sovrapposti di cellule poliedriche ancorate le une alle altre mediante i desmosomi. I cheratinociti di questo strato si assottigliano di spessore a mano a mano che si spostano verso la superficie. Il citoplasma è abbondante, pallido centralmente e ipercromico alla periferia, ricco di tonofìlamenti. Gli strati basale e spinoso costituiscono il corpo mucoso di Malpighi.
- Al di sopra dello strato spinoso si trova lo strato granuloso, composto da due o tre piani di cellule appiattite e infarcite di fini granuli di cheratoialina, una sostanza tingibile con ematossilina, carminio e colori basici. La sostanza costituente tali granuli è la filaggrina (una proteina aggregante i filamenti) la cui riduzione è riscontabile in alcuni casi di ittiosi e di dermatite atopica. Alla ME si osservano, nei gruppi cellulari superiori dello strato spinoso e nel granuloso, dei tipici corpi lamellari, definiti corpi di Odland. Lo strato lucido (uno o due strati di cellule), sovrastante lo strato granuloso, è composto da cheratinociti appiattiti con nucleo atrofico, ripieno di una sostanza chiara (eleidina), che si colora con rosso Congo e con colori acidi. Lo strato lucido è ben sviluppato e visibile istologicamente in sede palmo-plantare. Nel citoplasma delle cellule dello strato lucido, la ME evidenzia gli abbozzi delle fibre cheratiniche.
- Lo strato corneo è composto da più piani di cellule molto appiattite ed embricate fra di loro, prive di nucleo, con citoplasma omogeneo, eosinofilo e membrana ispessita (corneociti). Questi piani si sfaldano continuamente sulla superficie della cute, dando origine alle squame elementari. Il numero dei cheratinociti basali eguaglia quello dei cheratinociti desquamati, mantenendo quindi costante lo spessore epidermico. Il tempo necessario per la differenziazione da cellula basale a cellula cornea è, in media, di 3 settimane.
- Alla ME il citoplasma di queste cellule è ricolmo di fibre cheratiniche immerse in una sostanza amorfa. La parte dello strato corneo, in fase di sfaldamento, ossia già parzialmente distaccata della superficie cutanea, viene definita stratum disjunctum. In condizioni fisiologiche, la continua desquamazione cheratinocitaria avviene in modo apparentemente insensibile (si veda Immagine 01).
- Lo strato corneo, che funge da resistente barriera nei confronti delle aggressioni cutanee, possiede anche una ricca componente lipidica, tramite la quale modula il passaggio delle sostanze esterne attraverso la cute. I lipidi funzionalmente più importanti contenuti nelle membrane delle cellule dello strato corneo sono i ceramidi, che sono attivi nel prevenire l’aridità della superficie cutanea e nel regolare la perdita transcutanea d’acqua (transepidermal water loss). I ceramidi sono un gruppo strutturalmente eterogeneo di sfìngolipidi. Lo strato corneo ne contiene almeno nove differenti, due dei quali, ceramide A e B, sono legati covalentemente all’ involucrina (proteina prodotta dallo strato spinoso). La riduzione dei ceramidi dello strato corneo coinvolge la patogenesi di alcune malattie cutanee e, in primo luogo, la dermatite da contatto, l’ittiosi e la dermatite atopica.
Regolazione della proliferazione e differenziazione epidermica
- Il continuo rinnovamento dell’epidermide è da collegare a numerosi processi mirati al mantenimento di un numero relativamente costante di cellule.
- L’equilibrio è regolato grazie all’interazione tra cheratinociti adiacenti, cheratinociti dello strato basale e giunzione dermo-epidermica, tra giunzione dermo-epidermica e derma sottostante, oltre che dal processo di cheratinizzazione con cui le cellule basali si trasformano in corneociti. L’omeostasi epidermica è controllata da integrine, fattori di crescita, equilibrio tra proteasi e antiproteasi, citochine, retinoidi e vitamina D, ione calcio e attivazione dell’apoptosi.
- Integrine I recettori integrinici costituiscono una famiglia di più di venti diversi recettori transmembrana, eterodimeri di subunità α e β. Le integrine sono maggiormente espresse nei cheratinociti basali e localizzate nei tipici siti di adesione focale e a livello degli emidesmosomi. Sono predominanti α6β4, α2β1 e α3β1; le β1 sono concentrate tra un cheratinocita e l’altro, mentre la α6β4 è presente sulla superficie basale dei cheratinociti, associata agli emidesmosomi.
- Le integrine formano un legame e una via di comunicazione tra le molecole della matrice extracellulare, come il collagene e la fibronectina (α2β1 e α3β1), provvedono all’adesione tra cellule e MB e sono coinvolte nei processi di trasduzione del segnale. Esse determinano, inoltre, la polarità dei cheratinociti, dirigendo il processo di cheratinizzazione dallo strato basale allo strato corneo. L’ α6β4 ha un’alta affinità per la laminina 5, prodotta dai cheratinociti, ed è un’importante componente dei filamenti di ancoraggio della lamina lucida della MB.
- Fattori di crescita Numerosi fattori regolano la crescita e la proliferazione epidermica attraverso meccanismi autocrini e paracrini. L’EGF (Epidermal Growth Factor, fattore di crescita epidermico) e il TGF-α (Transforming Growth Factor α, fattore di crescita della trasformazione α) si legano alla stessa tirosina chinasi tramite recettori presenti sulle cellule epidermiche degli strati basale e spinoso, per stimolarne la proliferazione. Il TGF-α è prodotto dai cheratinociti, mentre l’EGF è prodotto da tutte le cellule epiteliali dell’organismo. Il KGF (Keratinocyte Growth Factor, fattore di crescita dei cheratinociti), prodotto dai fibroblasti, è un potente stimolatore della mitosi dei cheratinociti e degli epiteli. Altri fattori di crescita bloccano la sintesi del DNA e la mitosi dei cheratinociti e ne promuovono invece la differenziazione, come il TGF-β.
Equilibrio tra proteasi e antiproteasi
- La lisi dei desmosomi, che permette alle cellule superficiali di staccarsi liberamente e di produrre le squame elementari, è causata dall’attivazione di alcune sostanze, le proteasi appunto, che vengono inibite nella loro azione da altre sostanze, gli inibitori delle proteasi, che si disattivano solo al momento giusto che è diverso da sede a sede e che sono pH sensibile.
- Citochine Sostanze di derivazione cheratinocitica sono le interleuchine (IL)-1α, IL-6, IL-8 e il GM-CSF (Granulocyte Macrophage-Colony Stimulating Factor, fattore di stimolazione della colonia di granulociti e macrofagi). Queste citochine svolgono un ruolo nella normale regolazione epidermica, nei processi infiammatori e di riparazione delle ferite cutanee. In particolare l’IL-1α promuove la crescita di colture cheratinocitarie, le quali esprimono un recettore ad alta affinità proprio per l’IL-1.
- Retinoidi e Vit. D L’omeostasi epidermica dipende dall’interazione dei diversi ormoni e fattori che controllano il bilanciamento tra proliferazione, differenziazione e apoptosi dei cheratinociti. Uno di questi fattori è rappresentato dalla vitamina A e dai suoi derivati, i retinoidi. L’acido retinoico modula sia la mitosi dei cheratinociti sia la differenziazione delle cellule, epidermiche. Altro importante fattore di regolazione omeostasica dei cheratinociti è la vitamina D3, sintetizzata a livello cutaneo sotto lo stimolo della luce.
- Calcio Nell’epidermide esiste un gradiente dello ione calcio, la cui concentrazione aumenta passando dallo strato basale allo strato granuloso. Il calcio interviene nella formazione dei desmosomi e nella stimolazione della proliferazione dei cheratinociti; quest’ultimo effetto, però, è inibito da alte concentrazioni dello ione.
- Apoptosi Le funzioni dell’apoptosi, o morte cellulare programmata dei cheratinociti, si esplicano nella regolazione del numero di cellule e nella difesa dai danni subiti dalle cellule da parte di radiazioni (raggi UV, radiazioni ionizzanti), infezioni e trasformazione neoplastica.
Altri elementi cellulari dell’epidermide
- I cheratinociti costituiscono la maggior parte della massa epidermica. Nell’epidermide sono anche presenti melanociti, cellule di Langerhans, cellule di Merkel e rari linfociti T.
- Melanociti Sono cellule grandi, di forma poliedrica o stellare, con prolungamenti dendritici, con il corpo localizzato nello strato basale, interposte ogni 5-10 cheratinociti (nelle sedi fotoesposte, i melanociti sono più ravvicinati rispetto alle aree coperte) e sotto lo strato basale, ma comunque sopra la lamina densa. I melanociti cutanei sono presenti anche nel pelo, cioè a livello di matrice, midollare e corticale.
- Essi originano, come l’epidermide, dagli abbozzi della cresta neurale del foglietto ectodermico dell’embrione e sono preposti alla sintesi della melanina (melanogenesi), contenuta in grani (detti melanosomi) nel citoplasma. I melanosomi hanno struttura e grandezza geneticamente determinate e originano dal reticolo del Golgi come una vescicola circondata da una membrana unitaria (premelanosoma), evolvendo successivamente attraverso una serie di stadi morfologicamente definiti (I-IV).
- Nel I stadio i melanosomi hanno struttura irregolare e contengono piccole vescicole formate dall’invaginazione della membrana limitante esterna; nel II stadio divengono allungati e formano delle striature ordinate, che agiscono come supporto per la polimerizzazione della melanina che inizia nel III stadio, in cui, infatti, i melanosomi possiedono attività ossido-riduttasica e iniziano a diventare scuri con la sintesi di melanina; a mano a mano che il corpo cellulare del melanosoma diventa ricco di pigmento (IV stadio), esso si sposta lungo i prolungamenti dendritici dei melanociti in senso centrifugo, in modo da trasferire il pigmento ai cheratinociti vicini.
- I melanosomi coinvolti nella sintesi della eumelanina (marrone o nera) sono ellittici e internamente contengono la melanina disposta in lamelle concentriche; quelli che sintetizzano la feomelanina (rossa o gialla), più rappresentata nei soggetti rutili, hanno forma sferoidale e contengono la melanina in microvescicole.
- I melanociti sono cellule prive di tonofìlamenti e di desmosomi e si evidenziano con l’impregnazione argentica e con la DOPA (DihydrOxyPhenyWanine, diidrossifenilalanina)reazione.
- Il pigmento melanico viene trasferito dal melanocita, a circa trenta cheratinociti più vicini ed è poi lentamente eliminato per via transcutanea, seguendo il destino degli epiceliociti. Sono state proposte tre modalità di trasferimento della melanina, anche se l’esatto meccanismo rimane da chiarire. Il primo meccanismo prevede che il melanocita secerna il melanosoma nello spazio intercellulare e che questo venga poi endocitato dai cheratinociti; il secondo meccanismo di trasferimento propone la fusione diretta delle membrane cellulari di melanociti e cheratinociti, con passaggio dei melanosomi da una cellula all’ altra. Infine, il terzo meccanismo prevede che i prolungamenti dendritici dei melanociti lungo cui viaggiano i melanosomi siano fagocitati dai cheratinociti.
- Il pigmento, se arriva a livello dermico, viene assorbito dai melanofori (cellule macrofagiche) e viene drenato, per via linfatica, ai linfonodi.
- Melanogenesi La melanina origina nei melanosomi in seguito a un processo biologico irreversibile promosso dall’enzima tirosinasi, attivo sulla tirosina. Dal punto di vista biochimico, le tappe della sintesi melanica possono essere così sintetizzate: la tirosina, contenuta nei melanosomi, viene trasformata in DOPA dalla tirosinasi. La DOPA si trasforma, a sua volta, in dopachinone e, infine, in melanina granulare per azione della DOPA-ossidasi. La DOPA-reazione, utilizzata in laboratorio, sfrutta la proprietà del substrato melanocitario di attivarsi a contatto con la DOPA, producendo deposizione di pigmento melanico. È positiva soltanto nei melanociti attivi e non nei melanofori.
- La melanogenesi è controllata da fattori genetici e ormonali: ormoni ipofisari (MSH [Melanocyte Stimulating Hormone, ormone di stimolazione dei melanociti] e ACTH [Adrenocorticotropic Hormone, ormone adrenocorticotropo]), ormoni steroidei (estrogeni, cortisolo), tiroidei, epifisari (melatonina), oltre che da fattori tissutali (poliamine, nucleotidi ciclici ecc.) e cellulari (citochine). La pigmentazione cutanea, controllata geneticamente e di cui si conoscono almeno trenta varianti cromatiche, è un carattere etnico e non è condizionata dal numero di melanociti, ma dalla loro attività melanogenetica e dal grado e dalle modalità della dispersione dei granuli melanosomiali; nella cute scura i melanosomi hanno dimensione maggiore e una membrana più spessa.
- La pigmentazione melaninica riveste una funzione difensiva cutanea nei confronti delle radiazioni UV. A questo proposito, vengono distinti sei fototipi (classificazione di Fitzpatrick), con cui si valuta la reattività della cute nei confronti delle radiazioni solari. Il fototipo I (cute molto chiara) si scotta sempre e non si abbronza mai. Nel fototipo II, soggetto con capelli biondi o rossi e cute chiara, l’abbronzatura è minima e la scottatura solare è facile. Il fototipo III si abbronza dopo adeguata esposizione e si scotta più difficilmente. Il fototipo IV ha carnagione e capelli scuri e si scotta difficilmente. I fototipi V e VI corrispondono a etnie con cute iperpigmentata (alcuni orientali e alcuni sudamericani e i neri africani). Ovviamente esiste un continuum progressivo di fototipi, con gradi intermedi.
- Cellule di Langerhans Sono cellule immunocompetenti di origine midollare, di forma stellare e con prolungamenti dendritici che si insinuano tra i cheratinociti dello strato basale epidermico (2-4 % della massa cellulare basale) e dello strato spinoso e granuloso. Sono presenti anche nel derma, nelle mucose, nei linfonodi e nel timo. Le cellule di Langerhans non contengono tonofilamenti e desmosomi, sono DOPA-negative e argento-negative, ma si colorano con i sali d’oro. Contengono caratteristici granuli citoplasmatici a forma di racchetta (granuli di Birbek), osservabili alla ME, e specifici recettori di superficie delle cellule immunocompetenti (antigeni MHC di classe II, CD1a, CD4, S100 e il recettore ad alta affinità per le IgE). Le cellule di Langerhans sono le APC (Antigen Presenting Cells) con funzione di captazione e presentazione degli antigeni ai linfociti T.
- Cellule di Merkel Sono cellule chiare associate a un assone terminale per formare un recettore tattile e sono particolarmente numerose su polpastrelli, labbra, mucosa orale e guaina follicolare. Si trovano nello strato basale epidermico e aderiscono ai cheratinociti mediante desmosomi. Inoltre, contengono cheratine, granuli citoplasmatici sferici e producono diversi neuropeptidi, come la cromogranina A, l’enolasi neurone-specifica (NSE, Neuron-Specific Enolase) e la sinaptofisina. Tali cellule sono evidenziabili istologicamente mediante la reazione immunoistochimica con anticorpi monoclonali diretti contro l’NSE. Le cellule di Merkel sono stimolate dalla interazione dei cheratinociti, cui rispondono con la liberazione di trasmettitori chimici dai granuli in esse contenuti.
DERMA
- Il derma è intimamente connesso con l’epidermide sovrastante, di cui costituisce il sostegno meccanico e il substrato per gli scambi metabolici. Lo spessore varia tra 0,4 e 4 mm in rapporto alla sede anatomica.
- I suoi strati sono sovrapposti in maniera non nettamente differenziabile, come avviene nell’epidermide. Analogamente a tutte le strutture connettivali, il derma è composto da una sostanza fondamentale, fibre e cellule. In esso si distinguono il derma superficiale (o corpo papillare), il derma medio (o chorion) e il derma profondo o reticolare. Il derma superficiale si insinua verso l’alto con digitazioni multiple nell’epidermide (papille dermiche). I tratti di epidermide compresi tra le papille vengono chiamati creste interpapillari.
- Il derma papillare è particolarmente ricco di fibroblasti, che proliferano rapidamente rispetto al derma reticolare. Il connettivo costituente il derma superficiale è composto da fibre connettivali, elastiche e reticolari sottili, dirette prevalentemente in senso perpendicolare alle papille. Nel derma medio e reticolare i fasci delle fibre sono più spessi e hanno decorso parallelo o obliquo alla superficie cutanea. Il derma reticolare viene suddiviso in una porzione superiore e una profonda. La prima si differenzia dalla inferiore per la presenza di una popolazione cellulare ricca di fibroblasti, altre cellule connettivali e infiammatorie provenienti dal plesso vascolare subpapillare.
Componente fibrosa
- Le tre componenti fibrillari dermiche prodotte dai fibroblasti sono: fibre reticolari, collagene e fibre elastiche. Le fibre reticolari si evidenziano con l’impregnazione argentica sotto forma di fine trabecolato, sono espressione di una neogenesi connettivale e rappresentano la fase precollagenica.
- Le fibre collagene, eosinofile, ben rappresentate nel connettivo dermico, conferiscono consistenza strutturale. Le tappe della loro produzione comprendono la fase di formazione di tropocollagene (spessore delle fibrille di 50-150 À), la fase di procollagene e la produzione di fibre collagene mature (con spessore di 750-1000 A e periodicità di 640 À). Esistono diversi tipi di collagene; nella cute si trovano prevalentemente il collagene di tipo I nel derma reticolare, il collagene di tipo III nel derma papillare e quello di tipo di IV nella MB.
- Le fibre elastiche si colorano con l’orceina e sono costituite da fini fibrille immerse in una sostanza amorfa. Le fibrille non hanno struttura periodica e hanno spessore di 70 A. Le fibre sono immerse nella sostanza fondamentale dermica.
- Nel derma, inoltre, è presente la fibronectina, una glicoproteina che favorisce l’ancoraggio delle cellule alla matrice extracellulare.
Sostanza fondamentale
- È costituita da un gel contenente acqua, ioni, proteine, glucosio e mucopolisaccaridi acidi, di cui i più importanti sono l’acido condroitinsolforico, che regola il flusso elettrolitico attraverso la MB, e l’acido ialuronico, deputato all’idratazione dermica.
Componente cellulare
- Le varie cellule dermiche si differenziano tra loro in base a criteri di struttura, funzione, differenziazione e immunofenotipo.
- Le classi fondamentali di elementi cellulari abitanti il derma sono di origine mesenchimale (fibrociti, fibroblasti e mastociti) ed ematica (macrofagi, linfociti e cellule dendritiche).
Cellule di derivazione mesenchimale
- Fibrociti e fibroblasti Sono cellule di forma fusata e con nucleo ovalare, citoplasma abbondante e un voluminoso reticolo endoplasmatico che sintetizzano e degradano proteine della matrice connettivale e numerosi fattori solubili; inoltre, provvedono alla costituzione di un ‘impalcatura extracellulare, la cosiddetta “matrice”, con sintesi di laminina, fibronectina, vitronectina, trombospondina e collagene. Fattori solubili secreti da questa popolazione cellulare servono a favorire interazioni derma-epidermiche. La proliferazione dei fibroblasti è regolata da immunomediatori che includono IL-1α e IL-1β, IL-8 e HGF (Hepatocyte Growth Factor, fattore di crescita degli epatociti).
- Mastociti Sono cellule secretorie specializzate, caratterizzate da un citoplasma con granuli, presenti in grande quantità nel derma papillare vicino alla giunzione dermoepidermica, attorno ai vasi e alle diramazioni nervose del plesso subpapillare. I mastociti partecipano a reazioni di ipersensibilità di tipo immediato e sono coinvolti in malattie infiammatorie subacute e croniche. Queste cellule sintetizzano importanti mediatori, tutti contenuti in granuli preformati, tra cui istamina, eparina, carbossipeptidasi, fattore chemiotattico dei neutrofili e degli eosinofili. Inoltre, sintetizzano e rilasciano altre molecole, non contenute in granuli, tra cui numerosi fattori di crescita, citochine (IL-1, -3, -4, -5, GM-CSF e TNF-α), leucotrieni e fattori di aggregazione piastrinica. Tali mediatori attivano la permeabilità vascolare, con produzione di edema tissutale e reclutamento di cellule infiammatorie.
Cellule di derivazione ematica
- Cellule dendritiche Sono cellule stellate, analoghe alle cellule di Langerhans dell’epidermide, dotate di proprietà immunologiche, in quanto presentano un antigene dopo averlo fagocitato. Esse sono particolarmente abbondanti nel derma papillare e nella parte superiore del derma reticolare.
- Macrofagi Derivano da precursori del midollo osseo che si differenziano in monociti nel sangue per migrare nel derma, dove subiscono un’ulteriore differenziazione. Le loro funzioni includono: fagocitosi, processazione e presentazione dell’antigene a cellule linfatiche immunocompetenti, attività micobicida, tumoricida, secernente (fattori di crescita e citochine) ed ematopoietica; inoltre, i macrofagi sono coinvolti nella coagulazione, nell’aterogenesi e nel rimodellamento tissutale.
APPARATO PILO-SEBACEO
- Pelo, follicolo pilare, muscolo erettore del pelo e ghiandola sebacea annessa costituiscono l’unità pilo-sebacea, importante struttura annessiale cutanea.
Pelo
- È una struttura filiforme presente su quasi tutta la superficie cutanea; sono privi di peli (glabri) le superfici palmari e plantari, le semimucose e il dorso delle falangi ungueali. Gran parte della cute è provvista di peli corti ed esili (peli del vello o lanugo). Le caratteristiche del pelo sono determinate geneticamente (numero, colore, aspetto) e condizionate dal sesso, dall’età e dalla sede anatomica.
- La parte aerea del pelo è il fusto, mentre la parte che affonda nella cute costituisce la radice del pelo alloggiata in un’invaginazione epidermica che la riveste (follicolo del pelo) (Immagine 02).
- Il follicolo sbuca, a livello della superficie cutanea, in corrispondenza dell’ostio follicolare e subito al di sotto di questo si svasa a imbuto (infundibolo) (Immagine 03), per poi restringersi (collo o istmo follicolare), dove sbuca il dotto escretore della ghiandola sebacea annessa al pelo. Il follicolo diviene quindi cilindrico fino al colletto del bulbo, si restringe di nuovo prima di rigonfiarsi a livello del bulbo pilare (Immagine 04), il quale accoglie alla sua base un’introflessione dermica (papilla del pelo) con le sue sottilissime terminazioni nervose e vasali (capillari papillari). La struttura del pelo è composta da una porzione interna, midollare, costituita da due o tre file di cellule cubiche scarsamente cheratinizzate, una porzione corticale con cellule appiattite, cheratinizzate, ricche di melanina, e una cuticola esterna, con strato di cellule cornee, appiattite, contenenti cheratina amorfa.
- La radice è circondata da una guaina epiteliale (guaina della radice), costituita da tre strati dall’interno all’esterno: cuticola della guaina, strato di Huxley e strato di Henle. La guaina della radice viene eliminata unitamente al pelo con la sua caduta (o muta) (si veda Immagine 04) Il follicolo è costituito dalla guaina follicolare epiteliale esterna e, perifericamente, da uno strato di connettivo (Immagine 05).
- A livello bulbare scompare la netta distinzione tra la radice e la sua guaina, mentre sono presenti cellule epiteliali germinative (matrice del pelo) in moltiplicazione attiva per la ricrescita della radice e della guaina del pelo.
- Le fasi di sviluppo pilare vengono definite: anagen (fase proliferativa), catagen (fase di regressione) e telogen (fase di riposo).
- La fase catagen inizia con il blocco della proliferazione della matrice del pelo, successiva scomparsa del bulbo e retrazione e assottigliamento della guaina epiteliale esterna. In seguito si attiva una nuova papilla follicolare in anagen, in prossimità delle cellule germinative del pelo in catagen. Il nuovo pelo con il nuovo follicolo spinge poi all’esterno il vecchio pelo espellendolo.
- Annesso al follicolo pilo-sebaceo è il muscolo erettore del pelo (muscolo liscio), che si inserisce in profondità a metà del follicolo; da qui con decorso obliquo si porta al di sotto della ghiandola sebacea e, in alto, al limite inferiore del derma papillare (si veda Immagine 02).
- Peli e follicoli sono impiantati obliquamente rispetto al piano cutaneo, con muscolo erettore e ghiandola sebacea localizzati in corrispondenza dello stesso lato verso il quale è inclinato il pelo. La contrazione del muscolo erettore del pelo determina la compressione della ghiandola sebacea e ne favorisce lo svuotamento, oltre a provocare l’erezione del pelo (orripilazione o “pelle d’oca”).
Ghiandole sebacee
- Sono ghiandole di tipo alveolare semplice o composto, costituite da lobuli che riversano il contenuto lipidico (sebo) in un dotto escretore il quale, a sua volta, si apre a livello del collo del follicolo pilare. Ghiandole sebacee non annesse al pelo sono localizzate alle labbra, al glande, alle piccole labbra e al prepuzio. La secrezione ghiandolare è di tipo olocrino, ossia le cellule secernenti vengono eliminate con il loro contenuto.
Ghiandole sudoripare
- Sono ghiandole tubolari semplici costituite da una parte secernente avvolta a gomitolo, situata profondamente nel derma o nell’ipoderma, e una parte escretrice (dotto escretore) di forma tubulare, il cui tratto intraepidermico costituisce l’acrosiringio e termina sulla superficie cutanea con il poro sudorifero.
- Dal punto di vista istologico, il tubulo secernente è costituito da uno strato esterno connettivale proprio, seguito da uno strato discontinuo di elementi epiteliali contrattili (cellule mioepiteliali) e, internamente, da una fila di cellule prismatiche secernenti. Il dotto escretore è composto da un doppio strato epiteliale.
- La maggior parte delle ghiandole sudoripare è di tipo eccrino e secerne sudore, che contiene il 98% di acqua e il 2% di composti organici (urea, creatina, acidi grassi), sali (cloruro di sodio), acido urico, acido lattico e ammoniaca. Tali sostanze, unitamente al sebo e ai lipidi di origine cheratinocitaria, costituiscono il mantello o film idrolipidico cutaneo. In sedi particolari, soprattutto sessuali (regione ano-genitale, capezzolo), oltre ad ascelle, palpebre e condotto uditivo esterno, sono presenti ghiandole sudorifere apocrine, la cui secrezione è di tipo merocrino (eliminazione di propaggini citoplasmatiche cellulari sporgenti verso il lume canalicolare). Le ghiandole apocrine hanno anche peculiari caratteristiche anatomiche: maggiori dimensioni rispetto alle ghiandole eccrine, localizzazione più profonda del loro gomitolo secernente, sbocco del dotto escretore nell’infundibolo pilare.
- Il sudore apocrino ha odore caratteristico in ogni individuo. La natura e la funzione delle ghiandole apocrine, infatti, si ricollegano filogeneticamente al significato delle strutture correlate a funzione odorosa di richiamo sessuale (ferormoni).
Unghie
- Sono lamine di origine epiteliale, fortemente cheratinizzate, site sulla superficie dorsale delle falangi distali, dure, convesse in senso longitudinale e trasversale.
- La lamina ha una superficie dorsale e una ventrale o palmare, a contatto con il letto ungueale. La porzione prossimale del letto ungueale si continua con la matrice ungueale, sede germinativa della lamina.
- La lamina ungueale è composta da un corpo, due margini laterali, un margine distale e una radice. Quest’ultima è inserita nel vallo ungueale, una piega epidermica che dalla superficie dorsale delle falangi distali si introflette a ricoprire la radice. Tale plicatura cutanea viene detta eponichio nella porzione sovrastante la matrice, mentre due altre plicature costituiscono il perionichio, ai due lati della lamina.
- L’eponichio rappresenta la sede ideale per lo studio dei capillari cutanei, in quanto è l’unica superficie corporea in cui i capillari sono disposti orizzontalmente, permettendo la completa visione dei poli arterioso e venoso e dell’ansa. Il capillaroscopio permette lo studio morfologico e funzionale dei capillari, importante sussidio diagnostico in molte affezioni cutanee e sistemiche (collagenopatie, malattie del microcircolo, diabete). La porzione distale del vallo ungueale è costituita da un sottile bordo ipercheratosico (cuticola). La porzione anteriore della matrice è arcuata, a convessità distale, e traspare, per qualche millimetro, attraverso la lamina ungueale, con colore biancastro (lunula). La struttura della lamina è costituita da cheratinociti notevolmente cheratinizzati, molto appiattiti, con piccoli nuclei picnotici.
- Il solco ungueale (iponichio) è sito tra il margine libero dell’unghia e il polpastrello.
Vasi cutanei
- L’irrorazione cutanea è garantita da un ricco sistema arterioso, venoso e linfatico. Oltre alle arterie e alle vene proprie cutanee, la vascolarizzazione è caratterizzata da arterie muscolo-cutanee che attraversano i piani fasciali per raggiungere l’ipoderma e il derma. Esistono due vaste reti cutanee arteriose, venose e linfatiche, orientate parallelamente alla superficie cutanea; la più superficiale è sottopapillare, mentre l’altra è sita nel derma reticolare. Rami verticali vascolari connettono le due reti.
- Nelle papille del derma e del pelo giungono le estreme diramazioni vasali costituite dalle anse capillari, con caratteristica forma a forcina. Esse sono ben visualizzabili con la capillaroscopia al vallo ungueale, dove assumono una disposizione orizzontale. In tutte le altre sedi corporee le anse sono perpendicolari alla superficie cutanea e, quindi, è possibile osservare solo la sommità dell’ansa del capillare.
- Glomi arteriolari (peculiari anastomosi artero-venose) sono presenti nel derma reticolare soprattutto a livello dei polpastrelli delle dita.
- Per quanto concerne il sistema linfatico, i vasi linfatici cutanei sono numerosi e importanti nella regolazione della pressione interstiziale attraverso le loro funzioni di riassorbimento e di drenaggio dei fluidi. Inoltre, svolgono un ruolo di clearance tissutale da cellule, proteine, lipidi, batteri e sostanze di degradazione. La loro distribuzione ricalca quella dei vasi venosi e arteriosi. I collettori linfatici cutanei drenano ai linfonodi tributari sottocutanei, con prevalente localizzazione in sedi inguinali, ascellari e cervicali.
- Il flusso linfatico è regolato dalla contrazione muscolare. La stasi della linfa nei tessuti è prevenuta da valvole specifiche bicuspidi presenti nei vasi linfatici.
NERVI CUTANEI
- Nella cute sono presenti fibre del sistema cerebro-spinale, prevalentemente sensitive, e fibre vegetative del sistema nervoso simpatico (adrenergiche e colinergiche).
- Le fibre cerebro-spinali si arborizzano fino a giungere al derma papillare con fine intreccio anche attorno ai follicoli piliferi e alle ghiandole e hanno terminazioni libere. I nervi sensitivi forniscono innervazione ai dermatomeri, caratterizzati da confini imprecisi e da una parziale innervazione overlap.
- Le terminazioni nervose libere sono particolarmente sviluppate nel derma papillare e costituiscono importanti recettori sensoriali. Tali sono le fibre penicillate (recettori a rapido adattamento sensibili a calore, dolore e tatto), localizzate a livello subepidermico della cute dotata di peluria, e le terminazioni nervose del derma papillare a livello degli orifizi follicolari, sensibili al freddo.
- Le terminazioni corpuscolate sono invece strutture costituite da un insieme di fibre sensitive circondate da un involucro connettivale. Nello strato sottocutaneo sono presenti i corpuscoli di Pacini (sensibilità tattile) e di Ruffini (calorica), mentre nel derma si trovano i corpuscoli di Meissner (tattili) e di Krause (freddo).
- Le fibre del sistema nervoso vegetativo si distribuiscono ai muscoli erettori del pelo e alle ghiandole sudoripare apocrine (fibre adrenergiche) ed eccrine (colinergiche).
IPODERMA (O SOTTOCUTE)
- È uno strato di natura mesenchimale di spessore vario a seconda della sede anatomica, della costituzione corporea della persona e del suo stato di nutrizione.
- Gli adipociti dell’ipoderma sono grandi cellule rotondeggianti con citoplasma ricolmo di lipidi, soprattutto di trigliceridi, che comprimono il nucleo perifericamente. Nelle colorazioni routinarie, gli adipociti si mostrano otticamente vuoti, a causa della fissazione istologica con solventi.
- Queste cellule sono strutturate in lobuli delimitati da setti di tessuto fibroso connettivale, i quali, oltre alla funzione di organizzazione, fungono da sostegno alla vascolarizzazione arteriosa, venosa e linfatica e all’innervazione. L’ipoderma svolge numerose altre funzioni tra cui l’isolamento termico del corpo, la fornitura di riserva energetica, la protezione della cute, consentendone la mobilità sulle sottostanti strutture, e, infine, di segnale di regolazione della massa adiposa corporea, attraverso la secrezione dell’ormone leptina da parte degli adipociti. L’ipoderma manca tipicamente in alcune regioni del corpo come le regioni palmoplantari e il capo.
FISIOLOGIA CUTANEA
- La cute a tutti gli effetti è un organo, poiché è costituita da tessuti diversi ed esercita numerose funzioni specifiche. Qui di seguito verranno prese in considerazione più in dettaglio le varie funzioni assolte dalla cute, cui si è già accennato nei paragrafi precedenti.
Cheratinizzazione
- Rappresenta la funzione fondamentale della cute ed è il processo attraverso cui i cheratinociti basali si trasformano gradualmente in cellule dello strato corneo. Questo processo avviene mediante la progressiva sintesi di tipi diversi di cheratine, che vanno gradualmente a costituire un involucro cellulare insolubile e lipoproteico. Le cheratine hanno struttura filamentosa differenziata nelle cellule dei vari strati dell’epidermide; i cheratinociti basali producono le cheratine 5 e 14, quelli dello strato spinoso le cheratine 1 e 10. Alcune cheratine sono espresse soprattutto in alcune sedi: la cheratina 9, per esempio, è presente soprattutto in regione palmoplantare. Negli strati granuloso e corneo, i filamenti cheratinici si aggregano tra loro grazie alla filaggrina, una sostanza contenuta nei granuli di cheratoialina. Nello strato spinoso viene prodotta l’involucrina, proteina insolubile che va a costituire l’involucro corneo endoplasmatico. L’aggregazione tra questo involucro e le fibre di cheratina avviene nello strato granuloso a opera della proteina loricrina. Le sostanze lipidiche che dei corpi di Odland, presenti nei cheratinociti degli strati granuloso, lucido e corneo, contribuiscono alla formazione del film idrolipidico di superficie.
Funzione protettiva
- L’epidermide, e in particolare lo scrato corneo, rappresenta l’ interfaccia con l’ambiente esterno e svolge importanti funzioni di proiezione e difesa nei confronti di molteplici fattori esogeni.
- L’ epidermide, infatti, è esposta a continui stimoli quali irradiazione solare, UV e infrarossa, variazioni di temperatura ambientale, inquinamento atmosfrico, microrganismi patogeni ,sostanze chimiche, farmaci applicati per via topica, forze meccaniche e fisiche e corrente elettrica (sia pur a bassi voltaggi). Il derma costituisce la sua struttura portante connettivale e concorre a regolarne il tropismo. L’ipoderma, con lo strato adiposo, ha funzione di riserva e isolante e tende a mantenere costante la temperatura corporea: inoltre, assorbe e ammortizza l’energia meccanica proveniente da urti esterni
- Le fibre nervose afferenti meccano-, termo- e chemiosensitive hanno un ruolo protettivo, fornendo informazioni sugli stimoli propiocettivi e traumatici.
- Oltre a comportarsi come mantello protettivo fisico, la pelle possiede numerose altre proprietà che concorrono alla funzione protettiva:
- mediazione dell’infiammazione attraverso alcune molecole, quali glutatione, ossidasi, catalasi, sistema del citocromo P-450, vitamine C ed E;
- presenza di heat shock proteins, in grado di mantenere l’omeostasi cutanea;
- filtrazione dei raggi UV mediante la melanina, l’acido transurocanico e i metaboliti delle vitamine C e D;
- impermeabilizzazione della cute;
- metabolismo degli xenobiotici per via enzimatica mediante processi di glucuronizzazione e idrossilazione;
- difesa antimicrobica con i lipidi di superficie e il film idrolipidico, le proteine leganti il ferro, il complemento e i peptidi antimicrobici.
- Va sottolineato come lo strato corneo si caratterizzi per un pH marcatamente acido (pH da 4 a 6), che ne influenza significativamente la funzione antimicrobica; infatti, alcuni organismi patogeni quali stafilococchi, streptococchi e Candida per proliferare attivamente necessitano di un pH neutro. A tal proposito, giova ricordare che il mantello idrolipidico è dotato di un sistema tampone per il mantenimento del suo pH fisiologico. La cute è anche in grado di far fronte a insulti di agenti biologici, in quanto possiede un proprio sistema immunitario. La funzione immunologica dell’epidermide è anche legata alle cellule di Langerhans. Esse processano gli antigeni penetrati nella pelle e migrano ai linfonodi che drenano la linfa dalla cute, dove li presentano ai linfociti T, attivandoli. I linfociti T acquisiscono quindi specificità e memoria di risposta e vanno incontro a espansione clonale. Un’ulteriore esposizione cutanea allo stesso antigene, assieme all’ attività chemiotattica esercitata dalle citochine, dai cheratinociti e dai fibroblasti, determina la capacità, da parte del linfocita, di ritornare esattamente nel luogo di attivazione (homing skin).
- La pelle, quindi, assolve due fondamentali funzioni protettive; da un lato provvede a contrastare agenti patogeni esogeni per preservare la sua integrità e, dall’altro, assicura l’omeostasi dell’intero organismo, favorendo lo sviluppo e l’amplificazione della risposta immunitaria locale e sistemica.
Funzione sensoriale
- Grazie alla presenza di terminazioni sensoriali nervose per la meccanorecezione, la termorecezione, la sensibilità dolorifica e tattile, la cute è il più vasto organo di senso del nostro organismo. La sua organizzazione sensoriale è assicurata dalla presenza di fibre nervose sensoriali mieliniche. Queste si dipartono da tronchi nervosi principali verso il derma profondo, per formare il plesso profondo; da qui originano nuove ramificazioni che, giunte a livello del derma superficiale, danno luogo al plesso superficiale. Le diramazioni terminali si distribuiscono diffusamente nel derma e, in alcuni casi, oltrepassano la giunzione derma-epidermica come fibre nude, non mielinate. Le unità afferenti non hanno funzione specifica e percepiscono solo un tipo di stimolo.
- La nocicezione viene espressa da meccanocettori ad alta soglia e da nocicettori polimodali. Nella cute sono presenti anche unità funzionali efferenti neurosecretorie, che condizionano l’induzione di effetti viscera-motori e infiammatori nei tessuti periferici.
- Nella psoriasi, il fenomeno di Kobner potrebbe essere condizionato dal rilascio di neuropeptidi infiammatori nella cute traumatizzata. I neuropeptidi, inoltre, modulano il normale processo di riparazione delle ferite cutanee a cominciare dalla proliferazione cellulare, dalla produzione di citochine e fattori di crescita e dalla neoangiogenesi. La neuropatia diabetica e le lesioni del midollo spinale condizionano ulcere croniche insensibili a ogni terapia, a causa del deficit sensitivo cutaneo. Quindi, è evidente che la funzione sensoriale della cute va al di là della semplice trasmissione degli impulsi sensitivi al sistema nervoso centrale, in quanto rappresenta un’importante componente del sistema omeostatico cutaneo; inoltre, tale funzione svolge un importante effetto trofico per l’integrità e la funzionalità tissutale.
Funzione secretoria
- La cute assume un ruolo importante nella regolazione del bilancio idroelettrolitico in virtù della sua funzione secretoria. Infatti, essa è preposta alla secrezione sebacea, indispensabile per la costituzione del mantello idrolipidico cutaneo con sintesi di glicogeno, mucopolisaccaridi, lipidi (acidi grassi, steroidi, squalene) e proteine. Inoltre, la pelle riesce a disperdere il calore attraverso la produzione di sudore e la sua evaporazione, processi che avvengono in rapporto alle caratteristiche individuali e quando la temperatura esterna supera i 36 °C. Attraverso le ghiandole sudoripare di tipo eccrino viene prodotto fino a 1 L al giorno di secrezione sudorale con la perspiratio insensibilis e fino a 15 L in seguito a intensa attività fisica. Le anomalie della sudorazione includono iperidrosi (sudorazione in eccesso), ipoidrosi (in difetto) e bromidrosi (sudorazione maleodorante).
Funzione metabolica
- La cute assolve alla funzione metabolica con la sintesi di glicogeno, mucopolisaccaridi, biosintesi lipidica (acidi grassi, steroidi, squalene) e proteica. Il 20-25 % del colesterolo totale del corpo è prodotto dalla pelle. La sintesi di steroli e acidi grassi avviene sia nel derma sia nell’epidermide. In modo particolare, a livello dello strato basale si verifica il 60-70% della produzione lipidica totale della cute, sotto il controllo ormonale (variazioni di concentrazione degli ormoni tiroidei, testosterone ed estrogeni).
Funzione pigmento-genetica
- È correlata alla produzione di melanina e all’attività foto-protettrice. La principale proprietà della melanina consiste nella capacità di assorbire le radiazioni UV (280-400 nm) e la protezione dai danni che esse arrecano al DNA. Peraltro, anche i prodotti intermedi della biosintesi della melanina possono essere nocivi.
- I chinoni derivanti dalle reazioni tirosinasiche (dopachinoni) sono citotossici e inducono la morte cellulare, quando si accumulano ad alti livelli. La melanina è fotoreattiva e favorisce, in risposta alle radiazioni UVA, la produzione di specie reattive dell’ossigeno dannose per le cellule cutanee.
- Un incremento di feomelanina e di prodotti intermedi della sua biosintesi induce un’aumentata rottura delle catene di DNA, in colture di melanociti irradiati con UVA prelevati da individui di pelle chiara, rispetto a melanociti provenienti da soggetti di pelle più scura. Ciò fa supporre che la maggiore incidenza di melanoma fotoindotto in individui con i capelli rossi e pelle chiara non sia legato solo alla ridotta fotoprotezione, ma anche a una mutagenesi incrementata dalla feomelanina e dai suoi intermedi. L’abbronzatura costituisce la naturale difesa contro gli effetti nocivi delle radiazioni elettromagnetiche. Le radiazioni solari che attivano i melanociti sono i raggi UV compresi nel range del non-visibile tra i 290 e i 400 nm. Essi rappresentano fattori mutageni e promuoventi la possibile trasformazione tumorale dei cheratinociti, inducendo danno cellulare, mutazioni degli oncogeni e promuovendo la proliferazione melanocitaria. L’abbronzatura si sviluppa in due fasi, la prima transitoria e la seconda più tardiva, ma stabile. La fase transitoria ha un ruolo poco chiaro, poiché essa provvede scarsamente alla protezione contro successivi danni solari. In questa fase gli UVA inducono foto-ossidazione della melanina preesistente, dei precursori della melanina e/o dei metaboliti della melanina. Nella fase tardiva i raggi UV attivano la proliferazione dei melanociti, la produzione di melanina e il suo trasferimento ai cheratinociti.
Funzione termoregolatrice
- La cute ha un ruolo importante nella termoregolazione, sia perché fornisce informazioni termiche specifiche, sia perché agisce come organo effettore per il controllo della perdita di calore corporeo. La cute quindi esplica la sua funzione di termoregolazione con due modalità:
- organo effettore (modula le perdite di calore);
- organo efferente (informa i centri nervosi della temperatura esterna).
- Tale attività si esplica in un intervallo di temperatura compreso tra i 15 e i 45 °C, oltre i quali la sensazione di calore e/o freddo viene percepita come stimolo nocivo; infatti, la cute è particolarmente sensibile agli insulti determinati dalle temperature estreme dell’ambiente.
- Risposte reattive vasodilatatorie locali si osservano quando la temperatura raggiunge i 40 °C e risposte vasocostrittive quando la temperatura scende al di sotto di 10 °C; in tal modo si ottiene una protezione dal calore o dal freddo.
- Gli esseri omeotermi, come l’homo sapiens, spendono notevoli risorse fisiologiche per mantenere la loro temperatura interna (intorno ai 37 °C per l’uomo), poiché questa variabile fisiochimica condiziona profondamente molti processi biologici, quali il cambio chimico-fisico conformazionale e la conseguente attività di enzimi, recettori, canali di membrana; più in generale, essa condiziona la velocità delle reazioni chimiche cellulari. Ciò è evidente in caso di febbre; infatti, ogni grado di temperatura corporea in più determina un incremento del fabbisogno calorico dell’organismo del 13 % .
- Il corpo umano si comporta nei riguardi della temperatura ambientale come un sistema a due compartimenti, uno più esterno, maggiormente suscettibile a variazioni di temperatura, e uno più interno, che comprende gli organi vitali del capo e del tronco, con temperatura più elevata e regolata in un intervallo estremamente ristretto. Fattori vari quali l’ora del giorno e le fasi del ciclo mestruale possono determinare un’oscillazione di 1 °C anche a livello della temperatura interna. Inoltre, la temperatura corporea può aumentare in caso di intenso esercizio fisico o di febbre.
- Neuroni termosensitivi e centri regolatori cerebrali e spinali regolano con precisione la temperatura, soprattutto a livello della parte centrale del nostro organismo. La cute, invece, non è provvista di una regolazione cosi precisa e si caratterizza per una temperatura fortemente influenza dall’ambiente esterno. In particolare, i recettori termici endogeni e cutanei trasmettono infromazioni attraverso fibre afferenti al cervello e soprattutto all’ipotalamo dove termina la maggior parte delle informazioni relative alla temperatura. Questo sistema termoregolatorio deve garantire che la temperatura interna subisca minime variazioni e, quindi, che le condizioni ambientali influenzino solo gli strati più periferici dell’organismo.
- Il calore è trasportato attraverso l’organismo con due modalità: conduzione attraverso i tessuti e convezione attraverso il sangue; l’energia termica si distribuisce in questo modo dai tessuti più caldi e interni a quelli più freddi ed esterni e poi alla cute. La termoregolazione è regolata anche attraverso una ricca rete di shunt arterovenosi cutanei, in particolare in corrispondenza di naso, padiglioni auricolari, labbra, mani e piedi.
- Inoltre, il sistema vascolare cutaneo è regolato da fibre nervose distinte per funzione: le fibre simpatiche adrenergiche deputate alla vasocostrizione (anche se una minore attività di queste fibre determina, al contrario, una vasodilatazione) e le fibre parasimpatiche, deputate alla vasodilatazione.
- Al freddo le arteriole vanno incontro a costrizione come pure le vene superficiali degli arti, determinando un minor flusso sanguigno a questi livelli e una deviazione del sangue soprattutto alle vene profonde degli arti, le quali possono così ricevere energia termica dalle adiacenti arterie profonde con la conseguente conservazione di calore.
- Un ulteriore meccanismo nel mantenimento dell’energia termica è rappresentato dalla piloerezione al freddo (fenomeno dell’orripilazione). La contrazione contemporanea di un elevatissimo numero di muscoli lisci erettori del pelo determina una reazione esotermica, con ripristino dell’omeostasi termica cutanea. Un controllo comportamentale consente all’uomo, attraverso gli indumenti, la sopravvivenza anche in condizioni climatiche estreme, mentre il controllo fisiologico regola i sofisticati processi di adeguamento della temperatura interna.
Funzione di assorbimento e di permeabilità selettiva
- Lo strato corneo si comporta come una membrana omogenea a permeabilità selettiva, che modula la penetrazione di acqua e di altre sostanze applicate alla superficie cutanea. La proprietà diffusiva varia da soggetto a soggetto, in base alla regione corporea (dorso, arti superiori, tronco/ arti inferiori, addome, fronte) e in funzione delle caratteristiche morfologiche della regione cutanea. Un’importante funzione è svolta dai lipidi sintetizzati nello strato corneo. La capacità di una sostanza di attraversare lo strato corneo dipende dalla sua solubilità e dalla sua diffusibilità nell’acqua e nelle lamelle lipidiche. I lipidi assolvono tale compito grazie alla loro localizzazione intercellulare, concentrazione percentuale, organizzazione lamellare, composizione idrofobica, corretto rapporto molecolare e struttura molecolare compatta.
- La via di penetrazione translipidica cutanea ha permesso di utilizzare l’assorbimento sistemico di farmaci topici applicati sulla cute come steroidi e retinoidi, ma anche estrogeni (cerotti che contengono estradiolo per terapia anticoncezionale e terapia ormonale sostitutiva in postmenopausa), nitroglicerina, scopolamina e clonidina. Altre due vie di penetrazione transcutanea sono rappresentate dalla via transfollicolare e da quella trans-sudoripara.
Immagine 01
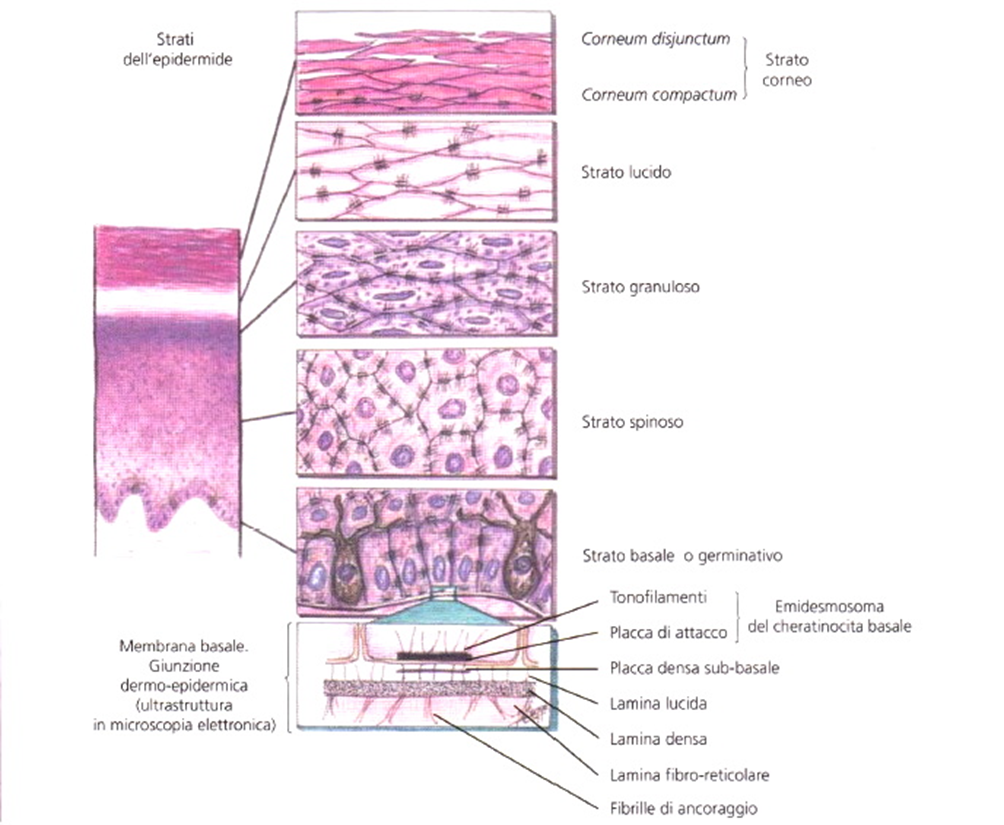
Immagine 01. Disegno degli strati dell’epidermide e della membrana basale.
Immagine 02
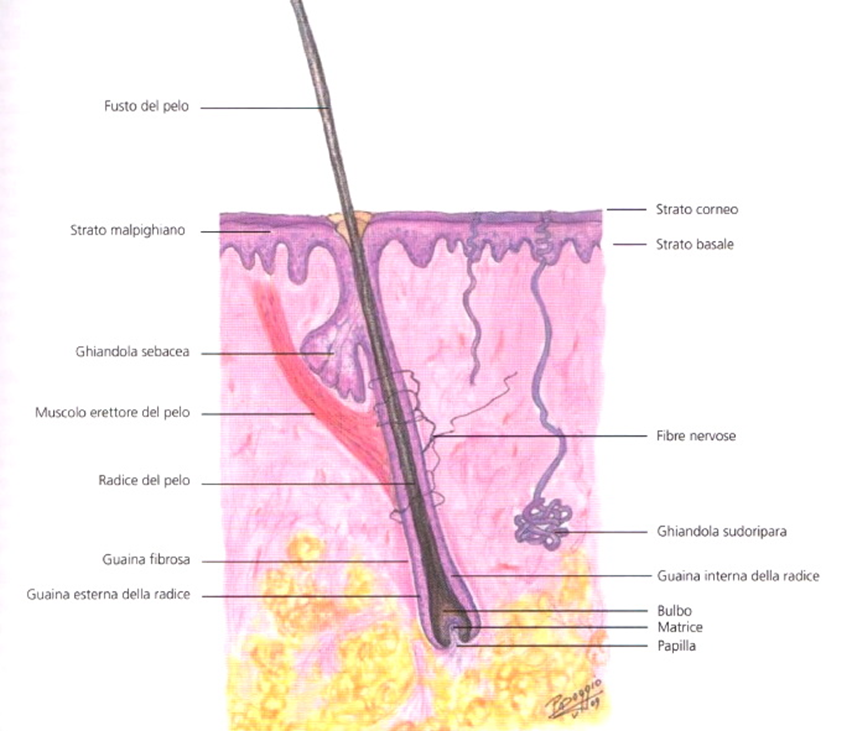
Immagine 02. Disegno del follicolo del pelo con annessa ghiandola sebacea e muscolo erettore del pelo. È mostrata anche la ghiandola sudori para eccrina.
Immagine 03
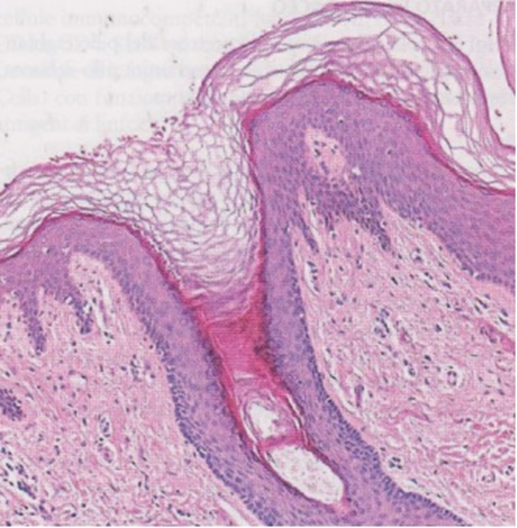
Immagine 03. lstologia della cute, con gli strati epidermici e la struttura del follicolo a livello infundibulare.
Immagine 04
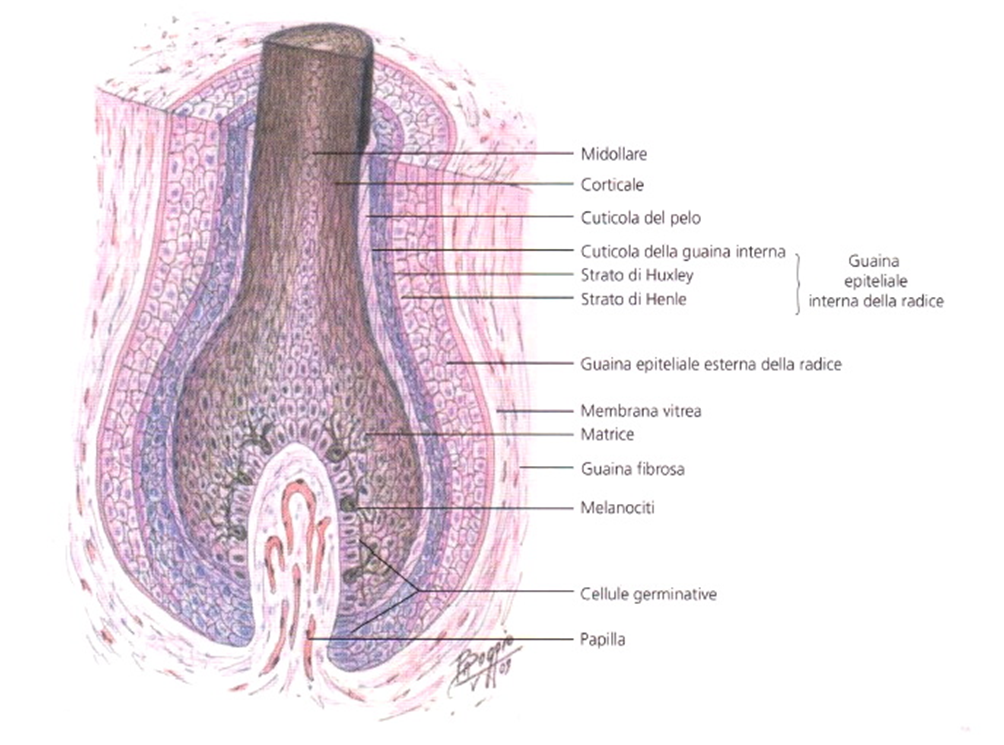
Immagine 04. Disegno del bulbo pila re con gli strati della radice.
Immagine 05
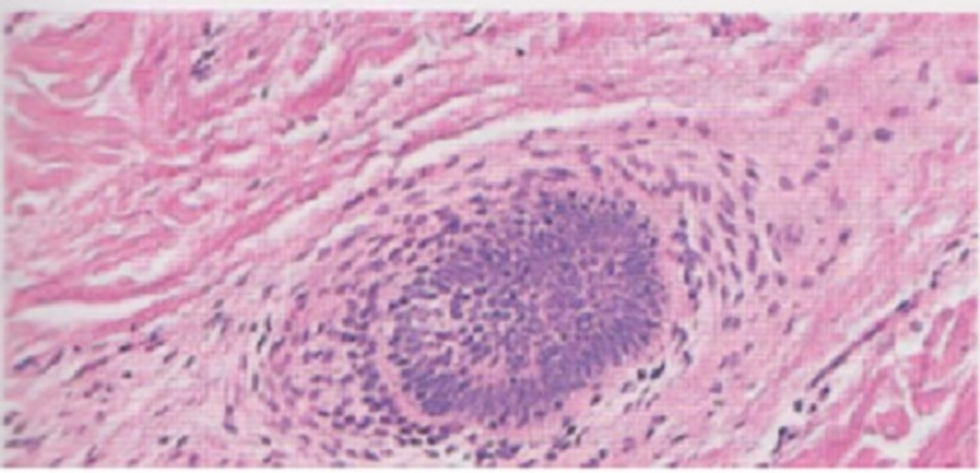
Immagine 05. Sezione trasversale del follicolo a livello della radice.